| |
Raw data: colon separated values (ASCII file)
| symbol |
parameter |
valori |
Refsori |
valpro |
Refspro |
LMM range |
literature range |
|---|
| pd |
daily mosquito survival |
Martens I scheme |
Martens 1997a |
Martens II scheme |
Martens 1997 |
Bayoh-12.5%, Martens I & II, Lindsay-Birley |
- |
| DgS |
standard Dg |
37 °days |
Detinova 1962c |
NU |
- |
standard/dry/humid Dg |
36.5-65.4 °days |
| TgS |
standard Ts |
9 °C |
Detinova 1962c |
NU |
- |
standard/dry/humid Ts |
4.5-9.9 °C |
| RH- |
Rainfall Humidity threshold |
NU |
- |
10 mm |
- |
- |
- |
| DgH |
humid Dg |
NU |
- |
37.1 °days |
Detinova 1962 |
- |
- |
| TgH |
humid Tg |
NU |
- |
7.7°C |
Detinova 1962c |
- |
- |
| DgL |
dry Dg |
NU |
- |
65.4 °days |
Detinova 1962c |
- |
- |
| TgL |
dry Tg |
NU |
- |
4.5°C |
Detinova 1962c |
- |
- |
| RR• |
rainfall laying multiplier |
1.0 |
Hoshen and Morse 2004 |
NU |
- |
- |
- |
| EGG |
number of produced mosquito eggs |
NU |
- |
80 |
cp. table |
50-200 eggs |
5-290 eggs |
| CAP |
cap to the number of egg laying mosquitoes |
NA |
- |
750 |
- |
100-2000 |
- |
| LR0 |
daily larval survival (lab) |
NU |
- |
0.825 |
cp. table |
0.80-0.95 |
0.82-0.99 |
| MMA |
Mosquito Mature Age |
14 days |
Jepson et al. 1948 |
12.0 days |
LR0- and pdl-table |
11.0-15.0 days |
11.2-27.6 days |
| Ds |
degree-days of the sporogonic cycle length |
111 °days |
Detinova 1962d |
111 °days |
Nikolaev 1935 |
111.0-148.0 degree-days |
111.0-204.4 degree-days |
| Ts |
temperature threshold of the sporogonic cycle |
18°C |
Detinova 1962d |
16°C |
Detinova 1962d |
14.0-18.0 |
14.2-19.0 |
| HBI |
Human Blood Index |
0.5 |
Hoshen and Morse 2004 |
0.8 |
cp. table |
0.5-0.9 |
0.0-1.0 |
| b |
mosquito-to-human transmission efficiency |
0.5 |
Hoshen and Morse 2004 |
0.3 |
cp. table |
0.1-0.5 |
0.01-0.50 |
| HIA |
Human Infectious Age |
14 days |
Hoshen and Morse 2004 |
20 days |
cp. table |
15-25 days |
12-30 days |
| r |
human recovery rate |
0.0284 |
Hoshen and Morse 2004 |
0.0050 |
e.g. Molineaux and Gramiccia 1980 |
0.0010-0.0150 |
0.0015-0.0385 |
| GF |
gametocyte fraction |
NU |
- |
0.5 |
cp. SAR in table |
0.3-0.7 |
0.1-0.7 |
| c |
human-to-mosquito transmission efficiency |
0.5 |
Hoshen and Morse 2004 |
0.2 |
cp. table |
0.1-0.3 |
0.0-0.379 |
| tri,h |
added infectious humans |
0.01 |
Hoshen and Morse 2004e |
0.00 |
- |
- |
- |
| tru,m |
added uninfected mosquitoes |
NA |
Hoshen and Morse 2004f |
0.00 |
- |
- |
- |
| tri,m |
added infectious mosquitoes |
0.00 |
Hoshen and Morse 2004 |
1.01 |
-g |
1.01 |
- |
Abbreviations
| symbol | symbol of the model parameter |
|---|
| parameter | model parameter |
|---|
| valori | value of the original LMM setting (Hoshen and Morse 2004) |
|---|
| Refsori | references with regard to the original setting |
|---|
| valpro | value of the proposed LMM setting |
|---|
| Refspro | references with regard to the proposed setting |
|---|
| Dg | degree-days of the gonotrophic cycle [°days] |
|---|
| Tg | temperature threshold of the gonotrophic cycle [°C] |
|---|
| pdl | daily larval survival (field) |
|---|
| SAR | Ratio between Sexual and Asexual parasite prevalence, i.e. the proportion of malaria parasite positive humans that are gametocytaemic [%] |
|---|
| NU | not used |
|---|
Comments
The fuzzy parameters with regard to the egg laying process and daily larval survival probabilities have yet not been specified (these are: FU0, FS1, FS2, FU2, LU0, LS1, LS2 and LU2).
The setting of LMM allows the usage of either per-gonotrophic cycle (α) or daily mosquito survival probabilities (pd).
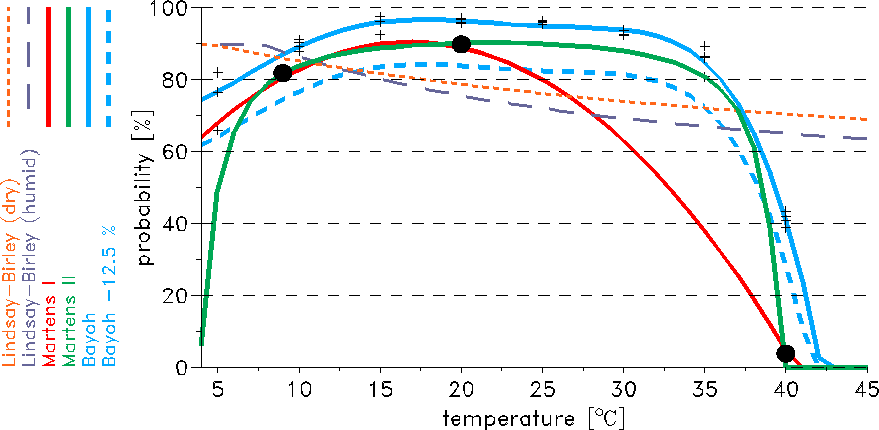
a Martens (1997) states: "Relying on data reported by Boyd (1949), Horsfall (1949), and Clements & Patterson (1981), we assume a daily survival probability (pd) of 0.82, 0.90 and 0.04 at temperatures (T) of 9°, 20° and 40°C, respectively, expressed as: pd=exp[-1.0/(-4.4+1.31 T-0.03 T2)]." However, Hoshen and Morse (2004) were not using the cited relationship, i.e. the so-called Martens II scheme. In fact they derived a different polygon from the three data points: pd=0.45+0.054 T-0.0016 T2, i.e. the so-called Martens I scheme.
b Bayoh (2004) observed the survival and mortality rates of Anopheles gambiae sensu stricto in environmental chambers at combinations of temperatures from 0 to 45°C at 5°C intervals and relative humidity's of 40%, 60%, 80%, and 100%. Using the data of these laboratory experiments and assuming an exponential model of mortality the following relationship has been computed: pd=-2.123 10-7 T5+1.951 10-5 T4-6.394 10-4 T3+8.217 10-3 T-2-1.865 10-2 T+7.238 10-1. However, vector survival is higher in captivity than in the wild (e.g. Clements and Paterson 1981) and hence pd has to be reduced. The shift-off was arbitrarily set to 12.5%. The probabilities of the Bayoh-12.5% scheme compare well with field observations. The so-called Bayoh-12.5% scheme agrees also fairly well with the mean daily survival of 84.6% found by Kiszewski et al. (2004).
c Shlenova (1938) performed experiments on the duration of blood digestion and ovarian development in Anopheles maculipennis. Various temperature and humidity combinations were used under controlled conditions. Detinova (1962) analysed this data set and found: "The sum of degree-hours at different humidities is composed of the differences between the actual temperature at each hour and the threshold temperature, which at a humidity of 30-40% is 4.5°C; at 70-80%, 9.9°C; and at 90-100%, 7.7°C. ... The difference between the mean diurnal temperature and the threshold temperature is calculated for each 24-hour period. These differences are added until the sum of the effective temperatures at 30-40% humidity is 65.4; at 70-80%, 36.5 and at 90-100%, 37.1."
d With regard to the temperature threshold of the sporogonic cycle Detinova (1962) firstly cites Nikolaev (1935): "Nikolaev (1935) established that the sporogony of Plasmodium vivax takes place at a temperature lower than 16°C, and of P. falciparum at a temperature of not less than 18°C."
Moreover, Detinova (1962) refers to the method of Moshkovsky (1946) for determining the duration of sporogony: "The sum of heat in degree-days for the maturation of the sporozoites in the mosquito's body is 105°C for P. vivax, 111°C for P. falciparum and 144°C for P. malariae. To establish the possible duration of sporogony it is necessary to subtract from the mean diurnal temperature in 24 hours the estimated lowest development temperature threshold, which is equivalent to 14.5°C for P. vivax and 16°C for other species of malaria parasites, and then in sequence to add the differences obtained in order to reach the total necessary for the development of the given species of parasite."
e Hoshen and Morse (2004) state: "The initial population is assumed to be non-infected, but new infections are being introduced at a constant low rate (trickle = 0.01 into a population of 100 every four days)."
f Hoshen and Morse (2004) state: "In the model, a small constant trickle (trickle2) of young uninfected mosquitoes (representing a new imported population) is added to the population of maturing mosquitoes."
g The interim LMM setting introduces the transmission of malaria by a constant trickle of infectious mosquitoes. Every ten days 1.01 new infectious mosquitoes have been added to the mosquito population. In this model version neither infectious humans nor uninfected mosquitoes are added.
|
|


{kind=link}